¿Cómo se produce la alergia? : las personas
alérgicas desarrollan anticuerpos (defensas)
de la clase IgE contra el veneno. En
posteriores picaduras (nuevas exposiciones al
veneno) la respuesta alérgica produce
síntomas, de gravedad variable.
¿Qué síntomas produce? : Las reacciones
alérgicas a picaduras suelen ser de tres tipos:
• Reacciones localizadas alrededor
de la picadura: con inflamación de la
zona (más de 10 cm de diámetro) y
dolor, generalmente persistentes más
de 24 horas. . No suele indicarse la
inmunoterapia. La mayoría de
reacciones locales requieren
exclusivamente la limpieza de la
herida (con agua y jabón) y la
aplicación de frío local (disminuye el
edema y actúa como analgésico). En
los casos más graves pueden
indicarse antihistamínicos y/o
corticoides.
• Reacciones generales (prevalencia
2.3% en población rural): urticaria,
edema, ahogo, bajadas de tensión
arterial, inestabilidad, hasta llegar a la
reacción anafiláctica y el choque, que
son los cuadros clínicos más graves
(1-5 muertes/10 millones de personas
/año).
• Reacciones retardadas: más raras y
en las que no se indica la
inmunoterapia.
¿Cuándo y como se producen? : los Polistes
suelen ser los responsables de la mayoría de
picaduras en piscinas, a obreros de la
construcción, electricistas, etc. La mayoría de
picaduras se producen durante los meses de
verano (mayor población de avispas y mayor
exposición de las personas), aunque no es
inhabitual observarlas fuera de esta estación.
Recomendaciones para el paciente
con alergia al veneno de
himenópteros.
En la mayoría de ocasiones los insectos pican
al sentirse amenazados. Evite:
• La proximidad de sus nidos, colmenas.
• Realizar aspavientos si alguno se le
acerca.
• Que sus familiares manipulen los
nidos.
Su agresividad aumenta con el calor y al
defender comida o bebida dulce
(especialmente refrescos), con los perfumes y
olores fuertes. Evite:
• Comer al aire libre
• Manipular contenedores de basuras.
• Realizar actividades de jardinería.
• Ir descalzo/a o con sandalias.
• Colonias, perfumes, ambientadores,
etc.
• Ropas holgadas y de colores vivos.
• Dejar la ropa al aire libre (en todo
caso, debe sacudirla antes de usarla).
Antes de iniciar la conducción de un vehículo
asegúrese de que no hay insectos en el
mismo, conduzca con las ventanillas cerradas.
En caso de una picadura por avispa:
• Incluso sin ser alérgico, se recomienda
no permanecer en la zona, ya que las
feromonas de alarma liberadas
durante el ataque podrían inducir
nuevas picaduras.
• Use la medicación que le haya
indicado su alergista • Acuda rápido al médico o servicio de
URGENCIAS más cercano.
Tras realizar el diagnóstico adecuado el
ALERGISTA valorará la necesidad de
tratamiento:
• INMEDIATO: generalmente con
ADRENALINA autoinyectable en
caso de reacciones graves. Retrasar
el uso de adrenalina utilizando otros
fármacos (corticoides,
antihistamínicos) puede provocar la
muerte del paciente. Pero la
adrenalina puede no ser
suficientemente efectiva en reacciones
muy graves, incluso el paciente puede
olvidar llevarla encima: por tanto este
fármaco no puede garantizar una
seguridad absoluta en caso de nuevas
picaduras.
• PREVENTIVO: mediante INMUNOTERAPIA
específica, único tratamiento
capaz de bloquear los síntomas en
caso de nuevas picaduras (de hecho
las dosis máximas suelen ser
equivalentes a dos picaduras).
viernes, 22 de julio de 2016



ENFERMEDADES TRANSMITIDAS POR LOS EXCREMENTOS DE RATAS
ENFERMEDADES TRANSMITIDAS POR LOS EXCREMENTOS DE RATAS .
La saliva y excrementos de las ratas y ratones transmiten las enfermedades más comunes.

Hantavirus: El virus, que se encuentra en la orina, la saliva y los excrementos del animal, se mezcla con la atmósfera por medio del aire o polvo. Los síntomas iniciales son dolores musculares y fiebre. Sin embargo, el síntoma primario de la enfermedad del hantavirus es la dificultad de respirar que es causada por la acumulación de fluidos en los pulmones. Principalmente los trasmiten los ratones de campo.

Los síntomas del hantavirus en el comienzo se confunden con pulmonía.

Leptospirosis: se transmite por la orina de las ratas , también con la heces y orina de perros y vacas. El cuadro clínico que se presenta es similar a una gripe con fiebre alta, escalofríos, sudoración, dolores de cabeza, dolores musculares y en la región lumbar. En algunos casos se pasa a una segunda fase con mayor gravedad se le denomina enfermedad de Weil y sus síntomas son manifestaciones renales tales como niveles altos de proteínas y presencia de sangre en la orina, lesión hepática que se manifiesta por ictericia.

· Teníais: Infestación del tubo digestivo por tenias, gusanos planos. Cada vez más infrecuentes, al procesarse correctamente las carnes para alimentación. Las tenias producen millones de huevos diariamente, que son evacuados con las heces. Las ratas los propagan por las heces y por animales infectados ingeridos por mamíferos. Cuando el hombre come carne de animales con gusanos, poco cocinada, se infecta. En la mayoría de los casos la infestación es sin síntomas.

· Triquinosis: Es una enfermedad parasitaria, producida por un nematodo denominado Trichinella spiralis. Transmitida por la rata por medio de los excrementos. Ataca a todos los mamíferos y puede ser transmitida al hombre. La infección ocurre generalmente por ingestión de larvas enquistadas en los músculos, éstas se liberan en el estómago y maduran en el intestino delgado. En las infecciones severas, que causan enfermedad y a veces muerte, los síntomas son: diarrea, náuseas, vómitos, fiebre moderada y dolor abdominal.
· Toxoplasmosis: es un término médico dado a una enfermedad infecciosa humana y de muchos otros animales, ocasionada por un parásito intracelular de distribución mundial, el Toxoplasma gondii. La enfermedad es considerada una zoonosis, es decir, existe normalmente en otros animales, pero puede ser transmitida a seres humanos. La fuente de infección primaria, son las heces de las ratas, la toxoplasmosis puede causar infecciones leves y sintomáticas, así como infecciones mortales que afectan mayormente al feto, recién nacidos, ancianos y personas vulnerables. Los síntomas más comunes son dolor de cabeza, dolores musculares, inflamación de los ganglios linfáticos, etc. En infecciones crónicas persiste como quiste en los tejidos, en otros casos menos comunes se presenta como enfermedad crónica.
jueves, 21 de julio de 2016
Entomología Básica orientado la familia Formicidae
Este pequeño curso está orientado a la familia Formicidae comentando por encima alguno de los aspectos exclusivos o no. Por ello antes de empezar con este curso sería recomendable que el lector leyera algún curso sobre entomología general.
Las secciones que tratare en este pequeño curso introductorio a la familia serán:
| 1.- Términos comunes 2.- Divisiones corporales 3.- La cabeza 4.- El mesosoma 5.- El pecíolo y post-pecíolo 6.- El gaster |
- Ácido poro (Acidopore): orificio de salida del ácido fórmico, característico de la subfamilia Formicinae. Está formado por el ápice del hipopigidio, algunas hormigas pueden ocultarlo tras el margen posterior del pigidio cuando no está siendo usado.
- Aguijón: parte del aparato defensivo presente en la mayoría de las subfamilias de hormigas.
- Ancho cefálico -AC- (Head width -HW-): es el ancho máximo de la cápsula cefálica medida en vista dorsal.
- Área frontal: una área triangular que está colocada posterior al clípeo.
- Articulación: área donde se juntan 2 secciones del cuerpo de una hormiga. Usualmente es móvil. Ejemplo: el área entre el pecíolo y el post-pecíolo.
- Bicarinado: con dos carinas, se refiere al clípeo. Usualmente hay dos dientes formados en el margen anterior del clípeo.
- Carina: Una cresta o borde en la superficie de la hormiga.
- Carina frontal: cresta que es parte de un par colocadas en la cabeza, que se extienden desde el clípeo hacia el lado posterior de la cabeza.
- Carina nuchal: La parte de la cabeza que separa las superficies dorsal y posterior.
- Clípeo: esclerito situado entre la frente y el labro. Es en este esclerito donde se articulan las mandíbulas.
- Dimorfismo: la existencia de obreras de dos tamaños distintos dentro de la misma especie.
- Distal: parte de un segmento más externo al cuerpo que otro.
- Dorsal: parte superior del insecto.
- Esclerito: placa dura que forma parte del exoesqueleto.
- Epinoto: térmico arcaico utilizado por los mimercólogos para designar el primer segmento abdominal. Actualmente se tiende a usar propodeo porque es el término universal utilizado en la morfología de los himenópteros.
- Epistomo: sinónimo a clípeo.
- Escapo (Scape): el primer segmento de la antena.
- Espina: estructura aguda en la superficie.
- Espiráculo (Spiracle): orificio del sistema traqueal que permite un intercambio de gases entre el interior y el exterior del cuerpo. En hormigas los espiráculos protoráxicos se han perdido encontrandose el primer par de espiráculos en el mesotórax. Los espiráculos propodeales son usualmente los más largos del cuerpo. En los segmentos abdominales 2-4 los espiráculos siempre son visibles, en los segmentos 5-7 frecuentemente están ocultos bajo el margen posterior de los tergitos precedentes y en el segmento octabo siempre está oculto.
- Estridulación: producción de sonidos mediante la fricción de dos partes del cuerpo. Parece ser una forma de comunicación que actúa cuando la química no funciona. (Las obreras de ciertas especies cuando quedan enterradas emiten sonidos detectables por las compañeras, las cuales las liberan).
- Fémur: tercer segmento de la pata, entre el trocante y la tibia. (Coxa-trocante-fémur-tibia-tarso).
- Feromona: sustancia química usada para comunicación entre individuos de una especie. Puede ser sexual, alarma, etc.
- Funículo: exceptuando al escapo, denominación que reciben el resto de segmentos de la antena.
- Gaster (gaster): morfologicamente se corresponde con los segmentos abdominales que van desde el tercer, o cuarto en caso de que exista post-peciolo, hasta el séptimo.
- Gena: sinónimo a mejilla.
- Haplodiploide: método de determinación sexual. El sexo esta determinado por el número de pares de cromosomas: el macho es haploide, la hembra es diploide. Ocurre en hormigas y otros himenópteros.
- Huevo trófico: huevo no funcional usado como fuente de comida en el nido. Producidos por la reina o por obreras.
- Esterno o esternóm: superficie inferior del segmento.
- Exoesqueleto: esqueleto o estructura de soporte externa producida por el ectodermo o epidermis y que no esta envuelto por tejido vivo. El exoesqueleto esta formado por numerosas placas denominadas escleritos.
- Larva: estado de crecimiento de los insectos con desarrollo holometabolo, le sigue el estado de pupa.
- Lóbulo: proceso redondo de las carinas frontales que puede cubrir las inserciones de las antenas, o también puede referirse a la esquina postero-lateral de la cabeza (lóbulo occipital).
- Longitud cefálica -LC- (Head Lenght -HL-): máxima longitud de la cabeza en vista dorsal y medida desde el borde anterior del clípeo hasta el borde posterior de los lóbulos occipitales.
- Masa: últimos segmentos de la antena de mayor tamaño que el resto.
- Mejilla: región de la cabeza colocada entre el ojo y la mandíbula.
- Mesosoma (alitrunk): es el segundo tagma reconocible en el cuerpo de las hormigas. Está formado por la fusión del verdadero tórax (pro-, meso- y metatórax) y el propodeo (primer segmento abdominal) formando una única unidad.
- Mesotórax (mesothorax): segundo segmento toráxico.
- Metamorfosis o desarrollo hemimetabola: tipo de desarrollo en el que el individuo sufre una serie de metamorfosis graduales hasta convertirse en adulto. Los estados son: huevo, ninfa y adulto
- Metamorfosis o desarrollo holometabola: tipo de desarrollo donde se diferencian un estado de crecimiento (larva), uno de diferenciación (pupa) y el reproductor (adulto). Los estados son: huevo, pupa y adulto
- Metatórax (metathorax): tercer segmento toráxico.
- Mirmecología: el estudio de las hormigas.
- Mirmecólogo: persona que estudia las hormigas.
- Monoginia: presencia de una sola reina en un nido.
- Monomorfismo: la existencia de obreras de un solo tamaño dentro de la misma especie.
- Nanítica: obrera muy pequeña, formada de la primera cria de la hembra cuando está estableciendo su nido.
- Ninfa: estado inmaduro (tras la eclosión del huevo) de los insectos con desarrollo hemimetabolo. El individuo pasara por una serie de metamorfosis graduales hasta convertirse en adulto.
- Nodo: sección del pecíolo que sobresale sobre la superficie.
- Ocelo: ojo simple cuya función básicamente consiste en medir la intensidad luminosa y en ningún caso puede llegar a formar imágenes.
- Occipital: región dorsal posterior de la cabeza.
- Ojo compuesto: ojo formado por numerosas unidades ópticas denominadas ommatidios.
- Omatidio: unidad del ojo compuesto.
- Palpo: proceso segmentado presente en la maxila y el labio.
- Pecíolo (Petiole): segundo segmento abdominal y usualmente separado del primer y tercer segmento abdominal por una constricción.
- Pectinada: se refiere a que presenta una fila de pelos muy finos.
- Pedúnculo: sección del pecíolo que conecta el gaster con el propodeo.
- Pigidio: el dorso del último segmento del gaster.
- Pleometrosis: formación de un nido inicial en que 2 o más reinas participan.
- Pleura o pleurones: superficies laterales del segmento.
- Polietismo: división de labores en miembros de la colonia, puede ser por casta o por edad. Cuando un individuo se especializa en varias labores durante su vida se hable de polietismo temporal. Normalmente las obreras trabajan con la cría al principio, después son guardias del nido y al final de su vida son forrajeras.
- Polimorfismo: la existencia de obreras de mas de dos tipos de tamaño dentro de la misma especie.
- Propodeo (propodeum): término universal utilizado en la morfología de los himenópteros y que hace referencia al primer segmento abdominal. Se encuentra fusionado al tórax.
- Protórax (prothorax): primer segmento toráxico.
- Pubescencia: pelos finos y cortos en la superficie.
- Pupa: estado de diferenciación e inactivo de los insectos con desarrollo holometabolo que sigue al estado larvario y precede al estado adulto.
- Queta: sinónimo a seda o pelo.
- Reticulada: se refiere a que el aspecto de la superficie es como una red.
- Sutura: línea o impresión formada por la unión de dos escleritos (placas corporales).
- Tarso (Tarsus, pl. tarsi): nombre genérico, y colectivo, que reciben los cinco segmentos apicales de cualquier pata. El primer tarso (primer tarsiómero) se articula con la tibia y recibe el nombre de basitarso (basitarsus), los tres tarsómeros siguientes no reciben ningún nombre en particular y el último tarsómero se denomina pretarso (pretarsus) y porta un par de uñas pretarsales (pretarsal claws).
- Tórax (Thorax): segundo tagma clásico en el cuerpo de los insectos. En hormigas, y en otros himenópteros, al verdadero tórax se le incorpora el propodeo luego el conjunto resultante no puede ser llamado como tal por ello se utiliza el término de mesosoma o alitrunk para referirse a este tagma en el caso de hormigas y otros himenópteros.
- Tergo (Tergite, = tergum, = tergal plate): superficie superior del segmento. Puede ser plano, curvado o estar especializado o subdividido en algunos segmentos. El tergo del protórax se denomina pronoto (pronotum), el del mesotórax mesonoto (mesonotum) y entre ambos se puede encontrar la sutura promesonotal (promesonotal suture) o bien estar formando un único esclerito llamado promesonoto (promesonotum); el tergo del metatórax se denomina (metanotum). A veces entre el mesonoto y el propodeo aparece el surco metanotal (metanotal groove) que es un surco o impresión transversa que representa el último vestigio del metatórax en la vista dorsal del mesosoma o alitrunk.
- Trofalaxis: intercambio bucal de alimento líquido entre miembros del nido.
- Vuelo nupcial: vuelo de apareamiento entre hembras y machos.
- Zángano: denominación que recibe el macho.
El cuerpo de las hormigas puede ser dividido, al igual que el de cualquier insecto, en tres partes sin embargo a diferencia del taxón al que pertenecen estas tres partes son cabeza, mesosoma y gaster. En otros grupos de insectos el mesosoma equivaldría al tórax y el gaster al abdomen, y de hecho a simple vista eso es lo que parecen, sin embargo el primer segmento abdominal denominado propodeo se encuentra incorporado al tórax, el segundo segmento abdominal se encuentra formando el pecíolo y, en caso de que exista post-pecíolo, este está formado por el tercer segmento abdominal.
| En las hormigas, al igual que en otro taxones, se ha producido una disminución en el número de segmentos abdominales permaneciendo constante el número de segmentos en la cabeza (seis más el acrón, componente no segmentario) y en el tórax (tres). En el abdomen tan solo se reconocen siete segmentos de los once iniciales existente durante las etapas iniciales del desarrollo embrionario. Según esto nos queda que el mesosoma esta constituido por cuatro segmentos: los tres propios del tórax más el propodeo, primer segmento abdominal; mientras que el gaster esta formado por seis o cinco segmentos, dependiendo de que aparezca o no el post-pecíolo. |  |
Es el resultado de la fusión de seis segmentos más un componente no segmentario denominado acrón. La cabeza de las hormigas es prognata, es decir, el eje longitudinal es aproximadamente horizontal al suelo por lo que las piezas bucales quedan orientadas hacia delante. Tiene gran importancia taxonómica y en ella se localizan la mayoría de órganos sensoriales así como el sistema nervioso central.
| La región dorsal de la cabeza, denominada epicráneo, esta marcada, generalmente, por una Y invertida denominada sutura epicraneal. La rama central, de la sutura epicraneal, se denomina sutura coronal y las dos ramas que parten de ella se denominan suturas frontales. Cuando se produce la muda, la vieja cutícula se rompe a nivel de estas suturas denominadas en su conjunto línea ecdisial y que corresponden a líneas de cutícula delgadas La parte de la cabeza que queda por encima de las suturas frontales se denomina vértex mientras que la que queda a ambos lados se denomina gena o mejilla. La porción de la cabeza encerrada por ambas suturas frontales se denomina frente. La frente puede estar delimitada por su zona distal, la más cercana al clípeo, por un surco epistomal que une las invaginaciones tentoriales anteriores (el tentorio es el nombre que recibe el endosqueleto de la cabeza). El clípeo se articula con el labro (una pieza bucal) por el surco clípeo-labral. Sobre él aparece un triángulo denominado triángulo frontal, además este clípeo puede presentar pubescencia. Detrás del vértex se encuentra el occipucio. A cada lado del clípeo se encuentran las articulaciones de las mandíbulas (piezas bucales). Detrás de cada mejilla aparece una invaginación tentorial posterior. Entre la cabeza y el tórax encontramos el cuello o cervix, la cual es una región flexible de origen intersegmentaria pero pudiendo incluir partes del segmento labial y del protórax, también se pueden encontrar parejas de escleritos cervicales. Los escleritos cervicales más importantes son los laterales que forman un fulcro entre la cabeza y el protórax, además es donde se insertan los músculos que provienen de la cabeza y el pronoto y que sirve para otorgar movilidad a la cabeza. |  |
Las antenas están formadas por una serie de segmentos el primero de ellos mucho mayor que el resto y denominado escapo, el resto de segmentos antenales se denominan funículos aunque el segundo segmento antenal también recibe la denominación de pedicelo. El número de segmentos antenales puede oscilar entre cuatro y trece. Los últimos segmentos antenales pueden aparecer engrosados formando la denominada masa antenal.
En taxonomía es muy importante el número de segmentos antenales (el escapo se incluye en la cuenta), la longitud del escapo respecto a determinados funículos antenales y en algunos casos también se tiene en cuenta la posición de la inserción antenal. El escapo se puede encontrar subdividido aparentando existir más de trece segmentos antenales.
Las piezas bucales que podemos encontrar son las mandíbulas (un par), maxilas (un par) y el labio (formado por la fusión de dos piezas semejantes a las maxilas). En el fondo de la cavidad bucal aparece un lóbulo no apendicular denominado hipofaringe y que esta sostenido por escleritos especializados. El labro también esta relacionado con las piezas bucales y a veces presenta receptores gustativos por su superficie interna.
| Las mandíbulas son las piezas bucales más importantes en las hormigas. Las hormigas al ser insectos ectognatos presentan mandíbulas con dos cóndilos (puntos de articulación). La cara interna de la mandíbula suele presentar un lóbulo incisivo distal dentado y un lóbulo molar basal. La relación entre estos dos lóbulos esta en función del tipo de alimentación, así las especies depredadoras presentan el lóbulo incisivo más desarrollado pues es mas importante agarrar la presa, para evitar que se escape, que la masticación. El lóbulo incisivo esta muy desarrollado en determinadas hormigas, hablándose de mandíbulas cepo o resorte. Los dientes se denominan dentículos y si aparecen grandes espacios entre ellos se denominan diastemas. Los dientes muy gastados o romos se denominan crenulados mientras que los largos y puntiagudos se denominan espiriformes. Cada maxila se compone de varias piezas, una basal denominada cardo y articulada con la cabeza por detrás de las mandíbulas y otra distal denominada estipe. El estipe porta dos enditos medios: el lateral llamado galea y el medio llamado lacinia; además porta un palpo maxilar lateral cuyo número de segmentos puede variar de uno a seis, siendo más frecuente encontrar seis. El labio esta formado por la fusión de las segundas maxilas entre si y cierra la cavidad preoral por la zona ventro-posterior. En el labio estructuralmente se pueden distinguir el postmentón y el prementón separados entre si por la sutura labial. El postmentón o postlabium es la región basal y fija del labio y se corresponde a los cardos maxilares fusionados, además puede subdividirse en submentón proximal y en mentón distal. El prementón o prelabium es la región distal y móvil del labio y se corresponde a los estipes maxilares fusionados, además presenta lateralmente un par de palpos (palpos labiales). Los palpos labiales, si aparecen, pueden estar formado por uno o cuatro segmentos, siendo lo más frecuente encontrar cuatro. En el labio podemos distinguir el postmentón, a veces dividido en submentón y mentón, que se corresponde a los cardos maxilares fusionados y el prementón, a veces bilobulado, que equivale a los estipes maxilares soldados. El prementón lleva un par de palpos laterales segmentados: los palpos labiales, compuestos por uno a cuatro artejos aunque lo más frecuente es que esta formado por tres. El borde apical del prementón suele presentar una prolongación a modo de lengua llamada lígula. La lígula típica esta formada por dos pares de lóbulos: las glosas, son los lóbulos internos y representan las lacinias maxilares, y las paraglosas, lóbulos externos que representan las galeas maxilares. Las glosas se encuentran fusionadas formando una ancha lengua. La formula palpar es el método estándar usado para indicar el numero de segmentos del palpo maxilar y el labial, así por ejemplo la fórmula palpar 6:4 indica que el palpo maxilar esta formado por 6 segmentos y el labial por 4. |
  |


Abarca desde la cabeza hasta el gaster y está formado por cuatro segmentos: protórax, mesotórax y metatórax-propodeo (este último se corresponde al primer segmento abdominal). Una de las características del orden Himenóptero, a excepción del grupo de los Symphyta, es la fusión del metatórax con el primer segmento abdominal o propodeo, al conjunto resultante se le denomina epinoto.
| El propodeo porta el primer par de espiráculos abdominales. El mesosoma en realidad es relativamente simple con un número limitado de placas y suturas salvo en las reinas donde existe mayor número de placas y suturas, esta complejidad adicional se requiere para la inserción de los músculos y el control de las alas durante el vuelo. La parte posterior del epinoto se denomina declive propodeal. Entre el pronoto y el mesonoto aparece la sutura promesonotal y entre el mesonoto y el epinoto la sutura mesopropodeal. En la pleura del epinoto, inmediatamente por encima de la coxa de la pata se encuentra la glándula metapleuralla cual a veces se sitúa en una depresión y puede estar cubierta o no por una escama cuticular. En la imagen se observa el orificio del primer par de espriculos respiratorios, marcados con el número 1; y la abertura de la glándula metapleural marcado con el número 2. Los únicos apéndices que podemos encontrar en las obreras son las patas. Las patas constan de un protopodio formado únicamente por la coxa y un telepodio, de tipo estenopodial (cilíndrico) y unirrameo (formado por una sola rama), y además consta de trocante, fémur, tibia, tarso y pretarso. La coxa es la base funcional de la pata, se encuentra articulada al tórax a través de la membrana articular denominada corial coxal y se articula con el trocante a través de dos cóndilos. El trocante es una pieza pequeña y dicondila, debido a esto último solo se puede mover en un plano. El fémur es la parte más fuerte y voluminosa de la pata. La tibia es una pieza esbelta y articulada al fémur a través de dos cóndilos. El tarso se subdivide en cinco pequeños segmentos denominados tarsómeros. El pretarso es la parte terminal de la pata. Apéndices toráxicos: las patas, como ya se comento anteriormente, presenta un protopodio y un telepodio de tipo estenopodial y unirrameo, además consta de cinco artejos: coxa, trocante, fémur, tibia y tarso. La coxa se articula con el tórax a nivel de la pleura o pleurón. El trocante une la coxa y el fémur, pero mientras que la unión coxa-trocante es móvil la unión trocante-fémur es fija; a veces el trocante parece dividido pero la pieza distal forma parte del fémur. En la unión tibia-tarso es común encontrar un espolón. |
|


La casta real además de los apéndices toráxicos ya comentados, las patas, presentan dos pares de alas. Las alas son expansiones tegumentarias aplanadas en forma de saco que surgen por evaginación del noto y la pleura del mesotórax y metatórax y se encuentran reforzadas por venas. La unión al cuerpo se realiza a través de un aparato apendicular diferente al de los verdaderos apéndices.
El pecíolo es un segmento, el segundo abdominal, que se sitúa entre el mesosoma y el gaster. Es una característica del subórden Apocrita, al cual pertenece la familia Formicidae (Órden Himenóptero ---> Subórden Apocrita ---> Superfamilia Vespoidea ---> Familia Formicidae). Está provisto de uno o más nodos y a veces su parte anterior se estrecha dando lugar al denominado pedúnculo. Puede estar seguido por un post-pecíolo o bien por el gaster. Pecíolo y post-pecíolo otorgan gran flexibilidad entre el mesosoma y el gaster.
Es la región posterior del cuerpo pudiendo estar formada por cuatro o cinco segmentos. El último segmento del gaster se denomina pigídio.
En los Acuelatas, grupo al que pertenecen las hormigas, siempre aparece un ovopositor aunque este no es empleado para la puesta sino que se transforma en un aparato de inyectar veneno: aguijón. Este aguijón consiste en un par de lancetas dentadas (valvas anteriores modificadas) y un estilete impar (valvas inferiores fusionadas), y que quedan enfundadas durante el reposo en un par de valvas dorsales blandas.
El aguijón se encuentra asociado a glándulas venenosas. La glándula venenosa es la denominada glándula ácida que es la más grande y dorsal de todas. Esta glándula ácida comunica con un reservorio de fuerte paredes musculares. Ventralmente se encuentra la glándula accesoria o alcalina también denominada glándula de Dufour y que actualmente no se considera parte del aparato venenoso, siendo relacionada con la lubricación del aguijón y secreción de sustancias de alarma o marcaje.
Cataglyphis ibericus
Cataglyphis ibericus es una hormiga termófila, muy veloz, que en los meses de verano forrajea en solitario a la búsqueda de cadáveres de insectos. Una de sus presas preferidas es la ubicua hormiga recolectora Messor barbarus, cuyos restos, una vez aprovechada la parte nutricia, son sacados y depositados alrededor de la entrada del hormiguero.

Desde hace tiempo vengo preguntándome si esta especie, además de carroñear, puede también cazar obreras vivas de Messor. Sin embargo, la actividad punta de las forrajeadoras de Cataglyphis se da en las horas cenitales de más calor, cuando las Messor y la mayoría del resto de especies se recogen en el interior de los nidos. A tales horas solo veremos a Cataglyphis recolectando infinidad de cadáveres.
Hace unos días ideé el siguiente experimento: capturé varias obreras vivas de Messor a las 11:00 (cuando todavía estaban activas) ,14:00 (cuando todas las Messor del entorno estaban ya bajo tierra por el calor reinante). La primera sorpresa que me llevé fue observar un comportamiento inmediato e inusual en las Messor: se subían rápidamente a los tallos cercanos, escalando hasta las puntas. No aguantaban la alta temperatura de la superficie del suelo.

Con la cámara del móvil en la mano, esperé a que alguna Cataglyphis localizara a alguna de las obreras de Messor encaramadas (las mismas Cataglyphis suelen subir a los tallos durante el forrajeo). En 10 minutos una de ellas localizó a una obrera Messor. El ataque fue fulminante. Por tres veces intentó cogerla y arrastrarla por las patas, aunque finalmente desistió.

En el siguiente video se aprecia el tercero de los ataques a la obrera de Messor:
https://youtu.be/A_Pt9KEG-Cs
Esta observación sugiere que, bajo ciertas condiciones, Cataglyphis ibericus puede predar sobre Messor barbarus. Probablemente, no en los largos senderos que transitan las Messor en sus horas de mayor actividad (cuando la temperatura es más benigna), pero sí, quizá, cuando comienza a hacer un calor sofocante y algunas obreras de Messor no han retornado todavía al nido.
Biología copulatoria del género Leptanilla
INTRODUCIÓN
En los últimos años ha cobrado un interés creciente el estudio de la biología copulatoria de las hormigas. Se investigan los fenómenos previos a la cópula, los mecanismos de acoplamiento, los procesos de transferencia espermática, la competencia entre espermas eyaculados, la selección críptica de la hembra mediante secreciones de su espermateca… Las técnicas moleculares permiten, además, conocer la composición genética de las colonias y la contribución respectiva de los sexuados. Todo ello resulta crucial para entender la evolución y organización de estos insectos sociales (Baer, 2010).
De manera general, dejando a un lado las excepciones, sabemos que hay especies monogínicas o poligínicas (con una o varias reinas fecundadas por colonia). También, que hay reinas monándricas o poliándricas (que copulan con un solo macho o con varios). La monandria implica un alto grado de parentesco entre los individuos de la colonia, lo que pudo haber dado origen al comportamiento altruista y, por ende, a la eusocialidad. Es por ello que se le considera un carácter ancestral de los himenópteros eusociales (Hughes et al., 2008; Boomsma, 2009). La poliandria, en cambio, sería un carácter derivado, generador de variabilidad genética intracolonial, y cuyo valor selectivo radicaría, entre otros factores, en una mayor plasticidad para la división de tareas, la adaptación a los cambios del medio o la evitación de infecciones, además de asegurar el llenado completo de la espermateca de las reinas (hipótesis de la limitación de esperma) (Crozier et al., 2001; Jaffé, 2014; Roes, 2015).
El CASO DE LEPTANILLA
El conocimiento detallado de la biología copulatoria de las hormigas se ha alcanzado, sin embargo, en solo unos pocos géneros y especies. Para el resto sigue siendo, en buena parte, terra incognita. Si entonces atendemos a géneros crípticos como Leptanilla, los problemas se multiplican. Cito a continuación algunas de las dificultades planteadas por estas hormigas minúsculas e hipogeas:
- Son escasas en las colecciones y difíciles de encontrar.
- Salvo algún caso excepcional, nunca se han colectado juntos machos y obreras, lo que ha dado lugar a una doble taxonomía pendiente de sinonimización.
- En la mayoría de especies se desconocen las reinas.
- Nunca se han observado sus cópulas.
¿Qué sabemos con razonable certeza del género Leptanilla? Que sus colonias son monogínicas (con entre 100 y 300 obreras), que son univoltinas (renuevan anualmente la población de obreras), y que comparten algunos de los rasgos del síndrome de las hormigas legionarias: nomadismo, reinas ápteras dichtadiiformes, probable fundación colonial dependiente mediante fisión de la colonia madre, y ciclos consecutivos y sincronizados de puesta de huevos (fase estacionaria) y eclosión de larvas (fase nomádica) (Masuko, 1990; Brady, 2003; kronauer, 2009).
HEBRA ESPERMÁTICA Y ESPERMATECA. VOLÚMENES SIMILARES QUE SUGIEREN MONANDRIA ESTRICTA
En relación con este tema de que vengo hablando, establecí recientemente una curiosa asociación entre dos elementos: a) las hebras espermáticas eyaculadas por varios machos vivos de Leptanilla SPA-02 que colecté en Madrid en 2015, y b) la espermateca que aparece en una micrografía de un corte sagital de una reina de Leptanilla japonica (Hölldobler et al., 1989).

El objeto inmediato de la asociación era saber si los volúmenes de la hebra espermática y de la espermateca eran similares o no. Dado que estaban implicadas dos especies diferentes –las únicas en las que hasta ahora se han podido asociar obreras y machos– tomé en consideración, a efectos de ajustes en la comparación, las longitudes totales de ambas castas de cada especie. L. japonica. Obrera: 1.2-1.3 mm (Baroni Urbani, 1977); macho: 1.3 mm (Ogata et al, 1995). L. SPA-02. Obrera: 1.5-1.6 mm ; macho: 1.5-1.6 mm .
Para medir el volumen de la espermateca de L. japonica la asimilé a una esfera y, utilizando la escala de la micrografía, apliqué la fórmula del volumen de la esfera. El resultado fue 0.000221 mm3.
Para hallar el volumen aproximado de la hebra espermática de Leptanilla SPA-02, la asimilé a un paralelepípedo rectangular. Estas hebras alargadas tienen estructura de cinta aplanada, no de cuerda con sección circular, tal como se aprecia en la siguiente fotografía:

La causa debe ser la rápida expulsión y solidificación del esperma al atravesar la falotrema, una estrecha hendidura longitudinal situada en el extremo del edeago:

Para medir la longitud (6.34 mm ) y anchura (0.02 mm ) de la hebra espermática, la aislé y monté cuidadosamente sobre un portaobjetos.

Para calcular el grosor de la hebra, procedí mediante el siguiente razonamiento: la hendidura (falotrema) que debe atravesar el esperma para salir al exterior, tiene una proporción anchura/longitud cercana a 1/10. Por tanto, el grosor de la hebra espermática vendría a ser, aproximadamente, la décima parte de su anchura (0.02/10). Teniendo ya la longitud, la anchura y el grosor de la hebra, apliqué la fórmula del volumen del paralelepípedo rectangular. El resultado fue 0.000254 mm3.
Ambos volúmenes, el de la espermateca de L. japonica (0.000221 mm3) y el de la hebra espermática de L. SPA-02 (0.000254 mm3), resultaron ser muy similares, con una diferencia que podría deberse, en parte, al distinto tamaño de las dos especies comparadas. Aunque se trata de una aproximación a groso modo, parece razonable concluir que el esperma de un solo macho de Leptanilla colma completamente la espermateca de la reina, impidiendo posteriores inseminaciones por otros machos. Ello sugiere que las reinas de Leptanilla son, muy probablemente, monándricas estrictas.
La monandria en Leptanilla concordaría con la ancestralidad asignada normalmente a este género, particularmente en algunas filogenias moleculares que sitúan a la subfamilia Leptanillinae como grupo hermano del resto de hormigas existentes (Brady et al., 2006; Moreau et al., 2006). Sin embargo, distaría en este carácter de las verdaderas hormigas legionarias (Dorylinae, Ecitoninae, Aenictinae), notoriamente poliándricas (Kronauer et al., 2007; Barth et al., 2014). Un caso similar a Leptanilla sería el del género Simopelta, de hábitos legionarios pero estrictamente monándrica (Kronauer et al., 2010).
Se desconoce el número de machos y reinas vírgenes que produce anualmente una colonia de Leptanilla. Excepcionalmente, pude colectar en una muestra de tierra 234 individuos de L. SPA-02, de los cuales 224 eran obreras y 7 machos, esto es, un 3% de la población. Incluso si se aumenta este porcentaje por posibles pérdidas y vuelos ya realizados, no parece probable que los machos superen el 10% de la población. Este número relativamente escaso de machos por colonia (tan distinto del altísimo ratio machos/reinas que se da en las hormigas legionarias) (Kronauer et al., 2007; Barth et al., 2014), quedaría compensado, de cara al encuentro entre sexuados, por la densidad y cercanía, casi contigüidad, de los nidos de Leptanilla en determinadas zonas, según he constatado en varios años de excavaciones sistemáticas. En cualquier caso, queda pendiente el problema de cómo localizan los machos alados a las reinas vírgenes ápteras y subterráneas. Si no existe endogamia dentro de los nidos, y los vuelos de los machos son adaptativos (como parecen indicar algunos mecanismos especiales del batido de sus alas, destinados a vencer la extrema viscosidad relativa del aire para insectos tan minúsculos), entonces los machos, tras emerger a la superficie y realizar sus vuelos, penetrarán de nuevo en la tierra a la búsqueda de reinas. Se hace difícil imaginar cómo consiguen tal proeza, y cuántos perecen en el intento, pero la monandria y el llenado completo de la espermateca asegurarían, mediante una sola cópula y para varios años, la capacidad reproductora de la reina.
¿ESPERMA FLUIDO O ESPERMATÓFORO?
Las sorprendentes hebras espermáticas de los machos de Leptanilla pueden aportar luz sobre la biología copulatoria del género. En mis experimentos, los machos las segregaban al ser depositados, todavía vivos, sobre la superficie del alcohol de un vial. He aquí algunas de sus características:
- Son muy largas, aproximadamente 3 o 4 veces la longitud del macho.
- Eran eyaculadas en menos de 10 segundos, durante los compulsivos movimientos premortem de los machos.
- Se endurecían o solidificaban (sin perder elasticidad) en el momento de ser eyaculadas.
La solidificación de las hebras, proceso en el que debe estar implicada la secreción viscosa procedente de las glándulas accesorias (secreción que sirve de medio envolvente y fluido a los espermatozoides), hace sospechar que el macho de Leptanilla transfiere a la reina un genuino espermatóforo, esto es, una estructura unitaria que funciona a modo de cápsula portadora de espermatozoides y que evitaría la pérdida de esperma durante el proceso de transferencia. Al penetrar en la espermateca, la hebra irá plegándose en su interior hasta formar un denso ovillo, como parece apreciarse en la micrografía de la espermateca de Leptanilla japonica que muestro al principio de este artículo.
La rapidez con la que el macho de Leptanilla eyacula la larga hebra espermática en condiciones experimentales (< 10 s), indica un poderoso mecanismo de bombeo. Es probable que en condiciones naturales la transferencia de la hebra a la espermateca de la reina sea directa, sin fase previa de almacenamiento en la bursa copulatrix, como sucede en el género Atta (Baer et al., 2006).
LA EXTREMA MINIATURIZACIÓN PRODUCE ÓRGANOS DESPROPORCIONADOS PARA PRESERVAR LA VIABILIDAD: EL CASO DE LOS HUEVOS, ESPERMATOZOIDES, ESPERMATECA Y GENITALIA DE LEPTANILLA
Desde hace tiempo se ha advertido que algunos órganos y estructuras de las minúsculas hormigas del género Leptanilla son excepcionalmente grandes en proporción a su cuerpo. Todo indica que existe un límite a la miniaturización a partir del cual dichas estructuras dejan de ser viables o adaptativas. Así, las obreras poseen una glándula de veneno hipertrofiada (necesaria, sin embargo, para contener el veneno suficiente con el que paralizar a los centípedos de que se alimentan), y las reinas albergan ganglios torácicos y espermatecas desproporcionadas (Hölldobler et al, 1989); los machos tienen, por otro lado, unas de las genitalias más grandes y singulares de la familia Formicidae (Bolton, 1990; Brady et al., 2005).
Un caso particularmente notorio de tamaño desproporcionado es mencionado por Masuko (1990): los huevos de Leptanilla japonica son de tamaño similar a los de Eciton hamatum (0.26 y 0.3 mm , respectivamente), pero una reina de E. hamatum es 10 veces más grande que una reina de L. japonica. La explicación más convincente es que una mayor miniaturización de los huevos los haría inviables. Pero para fecundar esos huevos desproporcionados de Leptanilla harían falta, a su vez, grandes espermatozoides, hipótesis expuesta por Hölldobler et al. (1989) para dar razón de la enorme espermateca de la reina de Leptanilla. Estas son sus palabras: “Si los espermatozoides de Leptanilla son también del tamaño de los de las hormigas grandes, cabe esperar entonces que la espermateca de la reina de Leptanilla sea desproporcionadamente grande”.
El hallazgo de las hebras espermáticas de los machos de Leptanilla me permitió realizar preparaciones microscópicas y visualizar, por primera vez, los espermatozoides filamentosos de este género.

Dado que los espermatozoides formaban una tupida red entrelazada, solo fue posible medir algunos segmentos que se distinguían nítidamente en la periferia de la hebra. La longitud de los segmentos más largos (varios de ellos con un extremo suelto) era de 45 µm; por tanto, es muy probable que la mayoría de espermatozoides completos superara esta medida.

A efectos comparativos, pongo a continuación una lista con algunas de las especies en las que se conoce la longitud de sus espermatozoides:
Leptanilla SPA-02…………..... ≥ 45 μm
Pseudomyrmex termitarius……53 μm (Moya et al., 2007)
Lasius pallitarsis…….............. 65 μm (Burnett et al., 2014)
Atta sexdens……………………..... 67 μm (Baer et al., 2009)
Solenopsis invicta……………..... 70 μm (Lino-Neto et al., 2002)
Acromyrmex echinatior……….. 83 μm (Baer et al., 2009)
Leptothorax gredleri.............100 μm (Oppelt et al., 2007)
Mycetosoritis hartmanni…….. 139 μm (Baer et al., 2009)
Atta cephalotes……………….... 150 μm (Baer et al., 2009)
Neoponera villosa……………... 175 μm (Barcellos et al., 2015)
Queda patente, dada la extrema miniaturización de Leptanilla en relación con las demás especies, que sus espermatozoides son proporcionalmente muy grandes, y que se necesita una enorme espermateca para albergarlos, tal como predecía la hipótesis de Hölldobler et al. (1989).
Como una imagen vale más que mil palabras, vea el lector la extraordinaria diferencia de tamaño entre un macho de Leptanilla SPA-02 y un macho de Solenopsis invicta. Las longitudes de sus espermatozoides respectivos difieren, sin embargo, relativamente poco: ≥ 45 μm en Leptanilla y 70 μm en Solenopsis.

Cabe conjeturar que el estudio futuro de los órganos sexuales de los machos de Leptanilla revele, también, un hiperdesarrollo del reservorio espermático y de las glándulas accesorias, así como un poderoso mecanismo de propulsión del esperma vinculado a las enormes genitalias de estas hormigas.
Referencias
- Baer B. 2010. The copulation biology of ants (Hymenoptera: Formicidae). Myrmecol. News 14:55-68.
- Baer B, Boomsma J. 2006. Mating biology of the leaf-cutting ants Atta colombica and A. cephalotes. J. Morphol. 267: 1165-1171.
- Baer B, Djikstra MB, Mueller UG, Nash DR, Boomsma J. 2009. Sperm length evolution in the fungus-growing ants. Behav. Ecol. 20: 38–45.
- Barcellos M, Martins L, Cossolin J, Serrão J, Delabie J, Lino-Neto J. 2015. Testes and Spermatozoa as Characters for Distinguishing Two Ant Species of the Genus Neoponera (Hymenoptera: Formicidae). Florida Entomologist 98(4):1254-1256.
- Baroni Urbani C. 1977. Materiali per una revisione della sottofamiglia Leptanillinae Emery (Hymenoptera: Formicidae). Entomologica Basiliensia 2: 427-488.
- Barth MB, Moritz RF, Kraus FB. 2014. The evolution of extreme polyandry in social insects: insights from army ants. PLoS One. 2014 Aug 21; 9(8):e105621.
- Bolton B. 1990. The higher classification of the ant subfamily Leptanillinae (Hymenoptera Formicidae). Systematic Entomology 15: 267-282.
- Boomsma J. 2009. Lifetime monogamy and the evolution of eusociality. Philosofical Transactions of the Royal Society B, 364(1533): 3191-3207.
- Brady SG. 2003. Evolution of the army ant syndrome: the origin and long-term evolutionary stasis of a complex of behavioral and reproductive adaptations. Proc. Natl. Acad. Sci. USA 100:6575-6579.
- Brady SG, Ward PS 2005. Morphological phylogeny of army ants and other dorylomorphs (Hymenoptera: Formicidae). Syst. Entomol. 30:593–618.
- Brady SG, Schultz TR, Fisher BL & Ward PS. 2006. Evaluating alternative hypotheses for the early evolution and diversification of ants. Proceedings of the National Academy of Sciences of the United States of America 103:
- Burnett WE, Heinze J. 2014. Sperm Bundles in the Seminal Vesicles of Sexually Mature Lasius Ant Males. PLoS One. 2014 Mar 26; 9(3):e93383.
- Crozier RH, Fjerdingstad EJ. 2001. Polyandry in social Hymenoptera -disunity in diversity? Ann. Zool. Fennici 38: 267-285.
- Hölldobler B, Palmer JM, Masuko K, Brown WL. 1989. New exocrine glands in the legionary ants of the genus Leptanilla (Hymenoptera: Formicidae: Leptanillinae). Zoomorphology 108: 255-262.
- Hughes WOH, Oldroyd BP, Beekman M. & Ratnieks FLW. 2008. Ancestral monogamy shows kin selection is key to the evolution of eusociality. Science 320 (5880):1213-1216.
- Jaffé R .2014. An updated guide to study polyandry in social insects. Sociobiology 61: 1-8.
- Kronauer D. 2009. Recent advances in army ant biology (Hymenoptera: Formicidae). Myrmecological News 12: 51-65.
- Kronauer D, Johnson RA, Boomsma J. 2007. The evolution of multiple mating in army ants. Evolution 61: 413-422.
- Kronauer D, O’Donnell S, Boomsma J, Pierce N. 2010. Strict monandry in the ponerine army ant genus Simopelta suggests that colony size and complexity drive mating system evolution in social insects. Molecular Ecology, vol. 20 (2): 420-428.
- Lino-Neto J, Dolder H. 2002. Sperm structure and ultrastructure of the fire ant Solenopsis invicta (Buren) (Hymenoptera, Formicidae). Tissue Cell 34: 124-128.
- Masuko K. 1990. Behavior and ecology of the enigmatic ant Leptanilla japonica Baroni Urbani (Hymenoptera: Formicidae: Leptanillinae). Insectes Sociaux 37: 31-57.
- Moreau CS, Bell CD, Vila R., Archibald SB & Pierce NE. 2006. Phylogeny of the ants: diversification in the age of angiosperms. Science 312: 101-104.
- Moya J, Manzini K, Lino-Neto J, Delabie J, Dolder H. 2007. Sperm ultrastructure of five species of the Neotropical ant genus Pseudomyrmex (Hymenoptera: Formicidae). Acta. Zool. 88: 181-187.
- Ogata K, Terayama M, Masuko K. 1995. The ant genus Leptanilla: discovery of the worker-associated male of L. japonica, and a description of a new species from Taiwan (Hymenoptera: Formicidae: Leptanillinae). Systematic Entomology 20: 27-34.
- Oppelt A, Heinze J. 2007. Dynamics of sperm transfer in the ant Leptothorax gredleri. Naturwissenschaften 94: 781-786.
- Roes FL. 2015. Queen Polyandry and the Evolution of Parasite Virulence. Entomol. Ornithol. Herpetol. 4: 146.
lunes, 2 de mayo de 2016
XI Congreso Ibérico de Mirmecología – TAXOMARA 2016
Sin faltar a su cita anual, la Asociación Ibérica de Mirmecología (AIM) y el Foro Lamarabunta convocan para este verano, en la ciudad de Murcia, el XI Congreso Ibérico de Mirmecología en el que expertos y aficionados compartirán durante varios días (del 13 al 15 de julio) ponencias, paneles, talleres de identificación y salidas al campo para muestrear hormigas.
El programa completo, y el modelo de inscripción, se encuentran en este enlace:
miércoles, 16 de marzo de 2016
Viaje a las fuentes
Journey to the fountains
Ya sabe el lector que desde hace unos pocos años ando interesado en las minúsculas –casi microscópicas– hormigas del género Leptanilla: ciegas, subterráneas, crípticas y consideradas entre las más primitivas de la extensa familia Formicidae.
Los mirmecólogos comentan desde hace un siglo que son muy pocos los que han podido verlas vivas o colectarlas. De hecho, están ausentes de muchas colecciones y se desconoce en gran parte su biología, distribución y diversidad. Bajo este comentario subyace la creencia de que estamos ante una rareza mirmecológica, ante unas hormigas escasas y excepcionalmente encontradas.
Esta creencia, sin embargo, debe desterrarse. Aplicando métodos específicos para recolectarlas (en zonas templadas o cálidas del hemisferio oriental), resultan ser abundantes, ubicuas y más ricas en especies de lo hasta ahora imaginado. Este cambio de perspectiva abre un campo sugestivo de investigación: podemos tener una razonable certidumbre de que nuevos muestreos –incluso en zonas ya prospectadas– darán lugar, con relativa facilidad, al descubrimiento de nuevas especies. Si, como pretendo sugerir, este grupo de hormigas constituye un caso de éxito evolutivo, habrá que conjugar varios aspectos:
- La ancestralidad de la subfamilia Leptanillinae, que de acuerdo con algunos estudios moleculares se situaría en la base del árbol filogenético de la familia Formicidae.
- Su amplia distribución: desde la región mediterránea al sur de África, desde Yemen a China, desde Japón a Filipinas, Malasia, Indonesia y Australia.
- El lentísimo sistema de expansión colonial bajo tierra, consistente probablemente en la fisión o separación de reinas ápteras recién fecundadas que apenas se alejarán unos pocos metros de la colonia madre.

En mis excavaciones en Madrid pude comprobar que las colonias se componen de 100 a 250 obreras, y que están cercanas, con tendencia a la aparición sucesiva de nidos contiguos de la misma especie. Por ejemplo, en un transecto de 30 metros excavado en 2013, 15 muestras de 20 kg de tierra contuvieron obreras de L. charonea (374 individuos en total), y 8 contuvieron obreras de L. SPA-02 (129); en otro transecto de 30 metros excavado en 2014, 16 muestras contuvieron obreras de L. SPA-02 (581), y solo 1 obreras de L. charonea (4).
Los machos –los únicos que salen al exterior, con alas y ojos bien desarrollados– realizan vuelos nupciales desde mediados de junio a finales de septiembre. En una piscina de 40 m2 situada a unos 50 metros de la zona de excavación pude recoger, en visitas diarias entre mediados de julio y finales de septiembre de 2013 y 2014, unos 900 machos de 4 especies diferentes. La recolección de machos flotando en piscinas, fuentes o estanques elimina el factor azar de las excavaciones, proporciona datos precisos sobre la diversidad y distribución de Leptanilla en cada zona, y asegura que en un radio cercano podrán encontrarse las obreras.
***
Adviértase que mi zona de búsqueda en Madrid se limitaba a media hectárea de terreno. Con la experiencia ganada y el convencimiento de que estas hormigas legendariamente raras debían ser notoriamente abundantes, decidí probar suerte en otros lugares fuera de Madrid capital. A tal efecto, en el verano de 2015 realicé cinco viajes de un solo día, y únicamente a centros urbanos a donde llegara el tren o el autobús. Visité San Lorenzo de El Escorial, Aranjuez, Guadalajara, Ciudad Real y Sevilla. Nada más llegar, por la mañana, recorría a pie la localidad buscando fuentes y estanques con agua en reposo. En los cinco sitios colecté un total de 87 machos de Leptanilla correspondientes a 6 especies diferentes. Por la tarde volvía a Madrid.

Tengo actualmente en mi colección siete tipos de machos de Leptanilla ibéricos diferentes, nombrados provisionalmente SPA-01, SPA-02… SPA-07. El único descrito formalmente es SPA-01 (Baroni Urbani, 1977). El macho SPA-05 me lo remitió desde Barcelona el profesor Xavier Espadaler. A la vista del pequeño muestreo realizado, cabe esperar nuevas sorpresas a medida que se exploren otras regiones.
Los machos son más fáciles de distinguir que las obreras, sobre todo por su genitalia. Algunas de sus estructuras –el edeago, los digiti y los telómeros– muestran claras diferencias entre las distintas morfoespecies. Muestro finalmente imágenes a 200 aumentos de las genitalias de estos siete machos ibéricos.

Referencias:
- BARONI URBANI C. 1977. Materiali per una revisione della sottofamiglia Leptanillinae Emery (Hymenoptera: Formicidae). Entomologica Basiliensia 2: 427-488.
Una nueva revista: NATURALISTA AFICIONADO
De la mano de José Alberto Fernández y un grupo de colaboradores expertos, nace hoy la revista online Naturalista Aficionado, un interesantísimo proyecto de Historia Natural abierto a todos.
El subtítulo de la revista define bien sus características: Ciencia y divulgación. Publicaciones de naturalistas aficionados, investigadores amateur y profesionales.

Desde aquí la enhorabuena a esta sugerente iniciativa, que a buen seguro constituirá un espacio vivo y acogedor de las observaciones y trabajos de muchos naturalistas, y un disfrute para todos.
Despliegue del arolium en Leptanilla
Unfolding of the arolium in Leptanilla
La contemplación y estudio de las estructuras biológicas, especialmente en especies cuya biología es poco conocida, resulta fascinante. La forma de dichas estructuras, producto de centenares de miles o millones de años de evolución, guarda los secretos de la interacción del animal con el medio.
Con estos pensamientos decidí inspeccionar el tarso de las primitivas, minúsculas y raras hormigas del género Leptanilla. Más concretamente quería saber si tenían arolium, una especie de almohadilla que poseen muchos insectos entre las uñas y cuya función es adherirse a las superficies lisas (donde las uñas resbalan y son ineficaces).
En 2014 había colectado en Madrid una colonia con 227 obreras de Leptanilla SPA-02 (pendiente de determinación) en la que aparecieron 7 machos. Este hallazgo me daba la oportunidad de buscar el arolium en las dos castas de una misma especie. Y pude confirmar que, efectivamente, tanto las obreras como los machos poseen arolia bien desarrollados y similares en los tres pares de patas. Extrañamente, Freeland et al (1982) no encontraron arolia en obreras de Leptanilla escheri, quizá porque pudieron haber utilizado ejemplares en seco. Keller (2011), en su revisión de la morfología de la familia Formicidae, sí afirma la existencia de arolia en los tres pares de patas de las obreras de Leptanilla swani, aunque no menciona su presencia o ausencia en los machos del género Leptanilla.
El arolium consiste en tejidos blandos y semitransparentes plegados de forma específica y situados en el pretarso. En el momento de aplicarse sobre el sustrato, el arolium se despliega aumentando considerablemente su volumen y la superficie de contacto. El mecanismo de despliegue parece ser hidráulico, probablemente por la presión ejercida por el líquido procedente de la glándula del arolium (Federle et al, 2001). Dos estructuras esclerotizadas en contacto con el arolium –el arcus (flexible) y el manubrium (rígido)– lo sostienen y participan en el proceso de despliegue.

Espoleado por los trabajos de Gorb (2001) y Gladun et al (2011) sobre la dinámica del arolium de los insectos, me propuse inducir artificialmente el despliegue del arolium de Leptanilla. Tras varias pruebas e intentos fallidos, pude contemplar finalmente el sorprendente proceso. El método empleado es sencillo: se separa primero una pata del cuerpo, en mi caso la pata trasera de un macho de Leptanilla SPA-02. La pata se sumerge a continuación en una micro gota de glicerina depositada sobre un portaobjetos, y se cubre con un cubreobjetos. Se coloca entonces el portaobjetos en la platina del microscopio (utilizando el objetivo de 40x) y, suavemente, se ejerce presión sobre el cubreobjetos mediante unas pinzas espatuladas.
Despliegue del arolium de la pata trasera de un macho de Leptanilla SPA-02. Vista dorsal
En estado natural el mecanismo hidráulico de despliegue debe ser muy rápido. La miniaturización de las estructuras del arolium en Leptanilla es extrema. Mientras en el estado plegado, en vista dorsal, la parte distal del arolium del macho de L. SPA-02 termina en punta, minimizando cualquier contacto con el sustrato, en su estado desplegado alcanza una anchura de 21 micras, aumentando la superficie total visible en más de dos veces.
Retornando ahora a la interpretación funcional de estas estructuras adherentes, debe resaltarse el hecho de que estén presentes –y bien desarrolladas– en hormigas estrictamente hipogeas que, en principio, parecerían no necesitarlas. En el caso de las obreras de Leptanilla –un género nomádico y tróficamente especializado–, es probable que participen en la sujeción de la cutícula lisa de los centípedos sobre los que predan, o que las empleen en las migraciones y el transporte de las grandes larvas. En cuanto a los machos, aparte de posibles funciones de sujeción de las hembras durante la cópula, hay que recordar que salen al exterior, que realizan vuelos nupciales entre mediados de junio y finales de setiembre y que, caídos sobre la superficie, deben buscar reinas en un periplo desconocido que quizá duré días o semanas. En este sentido, un arolium desarrollado podría serles de utilidad.
Curiosamente, varias especies de la subfamilia Amblyoponinae (que comparte con Leptanillinae los hábitos hipogeos y migratorios, la estenocefalia de las larvas, la predación sobre centípedos y la alimentación de la reina a base de hemolinfa larval) poseen también arolia, aunque a veces solo en las patas delanteras (Freeland et al, 1982). Un futuro estudio comparado de los arolia de diferentes especies hipogeas (su ausencia o presencia, y su morfología) podría iluminar algunos aspectos funcionales de estas notables estructuras adherentes y, por ende, la biología de estas crípticas hormigas.
Referencias
- Federle W., E. L. Brainerd, T. A. McMahon and B. Hölldobler. 2001. Biomechanics of the movable pretarsal adhesive organ in ants and bees. Proc. Nat. Acad. Sci. U.S.A 98 (11): 6215-6220.
- Freeland J., R. H. Crozier and J. Marc. 1982. On the occurrence of arolia in ant feet. Journal of the Australian Entomological Society 21: 257-262.
- Gladun D., S. N. Gorb and L. I. Frantsevich. 2009. Alternative tasks of the insect arolium with special reference to Hymenoptera. In Functional Surfaces in Biology, vol. 2, Adhesion Related Phenomena (ed. Gorb, S. N.): 67-104. Dordrecht: Springer.
- Gorb S. N. 2001. Attachment Devices of Insect Cuticle. Dordrecht: Kluwer Academic Publishers.
- Keller R. A.
Las hormigas en Cipriano de la Huerga (c. 1509-1560)
Cipriano de la Huerga fue un monje cisterciense nacido en León, catedrático de la Universidad Complutense y notabilísimo humanista y escriturista que contó entre sus discípulos a Fray Luis de León y Arias Montano. Buena parte de su extensa obra está perdida. En 1990 un grupo de estudiosos dirigidos por Gaspar Morocho inició la búsqueda, compilación y edición de todos los textos conocidos de Cipriano de la Huerga. Estas Obras Completas suman actualmente 10 volúmenes, publicados por el Secretariado de Publicaciones de la Universidad de León.
En el volumen VIII se incluye un curiosísimo texto titulado “Competencia de la hormiga con el hombre” (1559), descubierto entre los folios de un manuscrito de la Biblioteca del Palacio Real de Madrid. Lo editó y anotó Francisco Javier Fuente Fernández.

Primeras líneas del diálogo "Competencia de la hormiga con el hombre" de Cipriano de la Huerga, 1559. Manuscrito II-77 (3º) de la Biblioteca del Palacio Real de Madrid
En forma de diálogo entre dos hormigas, Cipriano de la Huerga afirmaba la superioridad del animal sobre el hombre. Con bello estilo y profundidad en los argumentos, recogió la tradición de los naturalistas grecolatinos Aristóteles, Plinio y Eliano.

Emblema utilizado por Cipriano de la Huerga en sus libros impresos
He seleccionado dos fragmentos del texto editado por Fuente Fernández, pero adaptándolo al español actual para facilitar su lectura.
Competencia de la hormiga con el hombre
(Fragmentos)
Cipriano de la Huerga (1559)
[…] El sentido del olfato en nosotras, hermana hormiga, es el que solo conoce la variedad de los sabores, porque en oliendo las cosas con diligencia las representa al que las ha de gustar y siente la fuerza de cada una de ellas, y sí toma solo aquello que es conforme a nuestra naturaleza, y lo que es contrario o impertinente valerosamente lo desecha no permitiendo que el gusto se estrague o se corrompa, y si alguna mezcla allí se halla de más, antes que la ponga delante al sentido del gusto la condena por su sentencia, pues de esta manera no somos fatigadas e importunadas de este sentido como lo son los hombres, los cuales, forzados de la variedad de apetitos insaciables que dije, mezclan juntamente el cinamomo, el bálsamo, el nardo, el incienso, el cálamo arábigo, la mosqueta, el ámbar, la algalia, conforme a los preceptos del arte médico ungüentario.
[…] Pues viniendo ya a lo que más hace a nuestro propósito, oso afirmar una cosa: que nunca entre los hombres hubo alguno de los que llaman filósofos, aunque en el ejercicio de las letras se haya aventajado mucho, que haya enseñado tantas leyes para bien vivir como nosotras. Crisipo, ni Crantor, ni Sócrates, ni Platón, enseñan mejor que nosotras lo justo, lo injusto, lo conveniente o dañoso. Nosotras entendemos los tiempos en los cuales se ha de buscar el mantenimiento necesario para la sustentación de la vida; llevamos a nuestros trojes grandes montones de grano; asentámoslos en nuestras casillas secretas con mucha prudencia; sabémoslo conservar para el invierno proveyendo que en un tiempo como este no sea necesario ir a buscar el mantenimiento por los campos vacíos y despojados ya de las mieses; nosotras llevamos con la boca tan grande peso que excede nuestra estatura y muchas veces nuestro poder, pero lo que falta de fuerzas remediamos con prudencia y con buen consejo y antes que escondamos las semillas para el tiempo del invierno las mordemos y cortamos de tal manera que no puedan de nuevo brotar de la tierra produciendo yerba o espigas, y, cuando por causa de las aguas han concebido demasiado humor, las sacamos a los tiempos al derredor de nuestras moradas tendiéndolas y revolviéndolas a una parte y a otra para que el calor del sol gaste la humedad superflua que podía ser dañosa al mantenimiento y por consiguiente a nuestra república, y todo esto hacemos con suma providencia teniendo siempre ojo al bien común.
Ni las noches sabemos estar ociosas mayormente cuando hay luna llena. De tal manera nos parece torpe y fea la ociosidad de suerte que ningún tiempo conveniente al trabajo dejamos perecer sin provecho, y cuando la luna esconde sus rayos cesamos del trabajo por parecernos que las tinieblas no son convenientes para nuestro ejercicio. Si algunas en el camino, trayendo a cuestas el grave peso, se sienten fatigadas, suceden otras de nuevo que han descansado para este fin, y si en el camino topamos otras que son de alguna ciudad vecina a la nuestra, aunque las favorecemos y ayudamos, pero tenemos mucha más cuenta con las que son de nuestra república y allí reconocemos nuestras banderas, y, como los que suelen apartar ejércitos, ordenamos nuestros escuadrones dando a cada uno cargo de llevar cierta cantidad de grano y aún, como tú sabes muy bien, todas las veces que nos encontramos en el camino nos saludamos unas a otras usando de buen comedimiento y de oficio, de caridad, preguntando si hay necesidad de nuestro favor, y en esta salutación nos detenemos tanto tiempo cuanto basta para saber las unas las necesidades de las otras y si es menester ayuda la damos con toda voluntad y a las de nuestra misma ciudad acariciamos y regalamos viendo qué es menester o por causa del largo camino o por otro accidente cualquiera. Lo contrario de esto habrás visto muchas veces entre los hombres, si con consideración lo has querido mirar.
Allende de esto, ninguna ciudad ni república fue jamás tan bien fundada ni tan puesta en orden por el saber de los hombres que pueda con razón cotejarse con la nuestra, porque como la experiencia lo enseña, ningún veneno es tan dañoso ni hay género de pestilencia tan pernicioso del bien común como es la ociosidad de los ciudadanos. Este mal está bien lejos de nosotras, porque ni nos espantan los calores del estío ni los rigurosos fríos del invierno, no los días ni las noches, para que dejemos de trabajar y enriquecer nuestra república de muchas maneras. Esto solo basta para entender cuán enemigas somos de la ociosidad, pues las noches que naturaleza dio a los otros animales para el reposo de los miembros fatigados, nosotras ordinariamente los empleamos en el acrecentamiento del bien común y no nos espanta la prolijidad del camino ni su aspereza, antes ordinariamente, siendo necesario traer el grano por lugares ásperos y pedregosos, no tanto con fuerzas cuanto con buena industria, llevamos nuestras cargas porfiando contra los más altos y más empinados montes que se nos suelen poner delante, y ansí, no con la grandeza del cuerpo ni con el vigor de los miembros, sino con la virtud del ánimo, sobrepujamos todas las dificultades, de donde parece haber sido más verdadera aquella sentencia que dijo una de las más antiguas de nuestro linaje: que ninguna cosa había puesto naturaleza en lugar tan alto ni tan difícil adonde la virtud y el valor de ánimo no pudiese llegar. De manera que no se puede dudar que, como la primera fuente de todos los vicios es la ociosidad, también la primera raíz de todas las virtudes es el ejercicio y el trabajo. A esta causa, tengo yo, hermana mía, presunción que somos más virtuosas, de mayor providencia y mayor justicia que el hombre, teniendo en tanta veneración el bien común y, según lo que él pide y aprueba, huyendo siempre el ocio y proveyendo a la necesaria sustentación de nuestra vida.
Pues si queremos descender en particular a las otras virtudes, ninguna se hallará estar desterrada de nuestra república, porque ¿quién no entiende que entre nosotras hay singular entendimiento y amor de la justicia? Nosotras tenemos determinados días para reconocer los pesos del mantenimiento que se ha traído, porque a cada uno de los escuadrones que dije, y a cada una de las familias por sí, se les da tanta parte cuanta es menester para su sustentación, lo cual, después de repartido, todos nuestros ciudadanos lícitamente y sin injuria de otro poseen, y nadie trata con su pensamiento usurpar lo ajeno ni tocar en la hacienda de su vecino, teniendo puesta siempre su confianza no sólo en la moderación del gasto y templanza, la cual siempre mora entre nosotras, mas también en la propia virtud e industria que siempre nos acompaña. No es de esta manera entre los hombres, los cuales, como vemos, gastando y destruyendo sus propios bienes pródigamente y sin juicio alguno ni parte de prudencia, comienzan luego a tratar con sus pensamientos como podrían vivir y sustentarse de la hacienda ajena, de donde nace que todas sus ciudades y repúblicas muchas veces las hemos visto ensuciadas no solo con extorsiones, con injusticias, contrarias, pero también con sangre derramada en las guerras civiles y con otros males innumerables, los cuales suele engendrar la discordia entre los ciudadanos.
¡Y qué diré yo de la templanza de las hormigas, la cual si se coteja con la del hombre es tanto mayor cuanto es mayor el cielo que la tierra! El gasto que entre nosotras se hace siempre es acompañado de prudencia, porque de tal manera remediamos a la necesidad presente que con la templanza en el comer proveemos a lo que está por venir, la cual virtud, aunque de suyo sea grande y admirable, pero debe ser a todos más agradable, porque es la propia guarda de la justicia, que si bien quieres mirar en ello la falta de esta virtud en la república humana es la que primero inventó ladrones, tiranos, homicidas, por la falta de esta virtud. Veras entre los hombres a unos presos, a otros echados del mundo con muertes infames y vergonzosas. Ninguna de estas cosas jamás acaece a nosotras las hormigas por el grande estudio y cuidado que tenemos en la guarda de la virtud, porque consideramos ser cosa digna de un buen ciudadano buscar siempre con grande cuidado y diligencia el bien común y acrecentarlo perpetuamente y conservarlo, pues no hay cosa más dañosa a la república de los hombres que anteponer el bien particular al provecho público de todos.
Ahora te ruego que juzgues con toda prudencia y cordura cuanto sea el hombre inferior a nosotras, pues era razón que supiese, tomando ejemplo de nosotras, nuestra manera de vivir, cuando constituyen alguna república, que la salud de los particulares depende de la salud pública y que según buen orden de naturaleza los ciudadanos deben con todas sus fuerzas defender la patria en la cual son nacidos y criados y enseñados de muchas maneras. Pero no hacen esto los hombres, sino antes todo lo contrario. Debían todo lo que aran y siembran y cogen referirlo al público provecho de la patria, cuya salud siempre ha de ser tenida por más digna y más antigua que la dignidad y salud propia, pues de esta manera las hormigas, pequeños animales, ponemos infinitos ejemplos de virtud delante los ojos del hombre soberbio y arrogante, a semejanza de las cuales, si la mayor parte de los hombres se quiere cotejar, ninguno se hallaría que sea dotado de tantas y tales virtudes, pues la mayor parte de ellos, viviendo licenciosamente, entregándose de todo punto a la ociosidad y deleite, de ninguna cosa viven cuidadosos tanto como de celebrar banquetes superfluos y demasiados, del beber hasta salir de juicio, de la superfluidad de los manjares y de los servicios de Venus y de su hijo, a los cuales tienen por dioses.
Sobre una hebra espermática en los machos de Leptanilla
Mientras manipulaba los machos de Leptanilla capturados este verano, advertí algo extraño. Los ejemplares estaban dentro de un pocillo con alcohol, y los intentaba girar y desplazar con la punta de un pelo de pincel enmangado en una varilla. Sorprendentemente, los movía sin tocarlos, como si el pelo de pincel se enganchara en algún objeto invisible asociado a los ejemplares.
Inmediatamente, puse debajo del pocillo una cartulina negra para ganar contraste, y aumenté el zoom de la lupa estereoscópica. Apareció entonces una larga hebra blanca que pendía del extremo de la genitalia y que se alargaba hasta cuatro veces la longitud del macho.

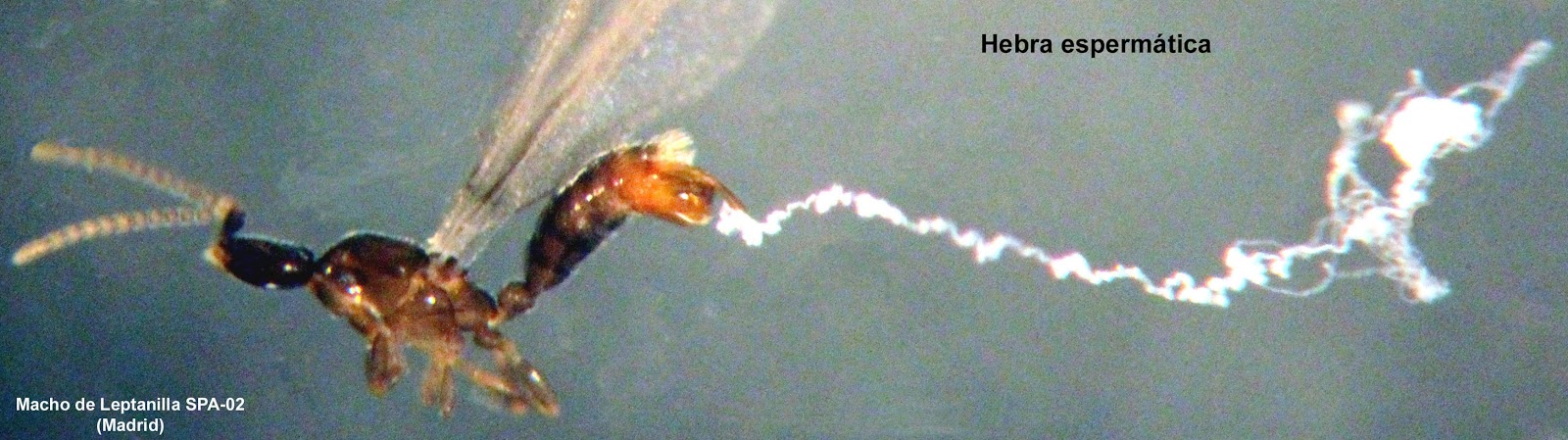
Para intentar averiguar de qué se trataba, realicé primeramente algunas preparaciones microscópicas introduciendo el macho con su hebra dentro de una gota de glicerina. Este medio de inclusión, que facilita la colocación estable del ejemplar en la posición deseada, me permitió ver exactamente el origen de la hebra, que surgía de la falotrema, una abertura con forma de ojal situada en el extremo de la cara dorsal del edeago o pene.


A 100 aumentos la hebra aparecía como un único y finísimo hilo con múltiples pliegues.

La hipótesis de trabajo era cada vez más evidente: podía tratarse del esperma de Leptanilla. En este punto, conviene detenerse un momento para preguntarse por las condiciones en las que se segregaron las hebras, que aparecieron casi exclusivamente en los machos colectados en 2015, y no en los colectados en 2013 y 2014. En estos dos últimos años recogí los machos flotando en piscinas, la mayoría de las veces muertos, y en todo caso metiéndolos en un frasco con agua para trasladarlos. En 2015, sin embargo, cambié de estrategia: coloqué trampas de captura consistentes en bandejas con agua en las que ponía una fuente de luz ultravioleta que se activaba de noche. Todas las mañanas acudía a las trampas y recogía el material que había caído. Buena parte de los machos seguían vivos, y los introducía directamente en un vial con alcohol al 70%. ¡Esta era la clave de la aparición de las hebras! Al depositar los machos todavía vivos sobre la superficie del alcohol del vial, realizaban movimientos compulsivos durante varios segundos antes de hundirse, periodo en el que segregaban las hebras.
Pero retomemos el relato de la indagación. La hipótesis de que se trataba de hebras espermáticas había que demostrarla. Dicho en pocas palabras: ¡tenía que ver los espermatozoides! Me puse a ello y comencé a hacer preparaciones microscópicas de las finísimas y blancas hebras de que disponía (previamente separadas, con mucho cuidado, del edeago de los machos). En una de las preparaciones correspondiente a un macho de Leptanilla SPA-02 (Madrid), sumergida la hebra en alcohol, teñida con azul de metileno y cubierta con cubreobjetos, creí distinguir a 400 aumentos una densa maraña de miles de espermatozoides.
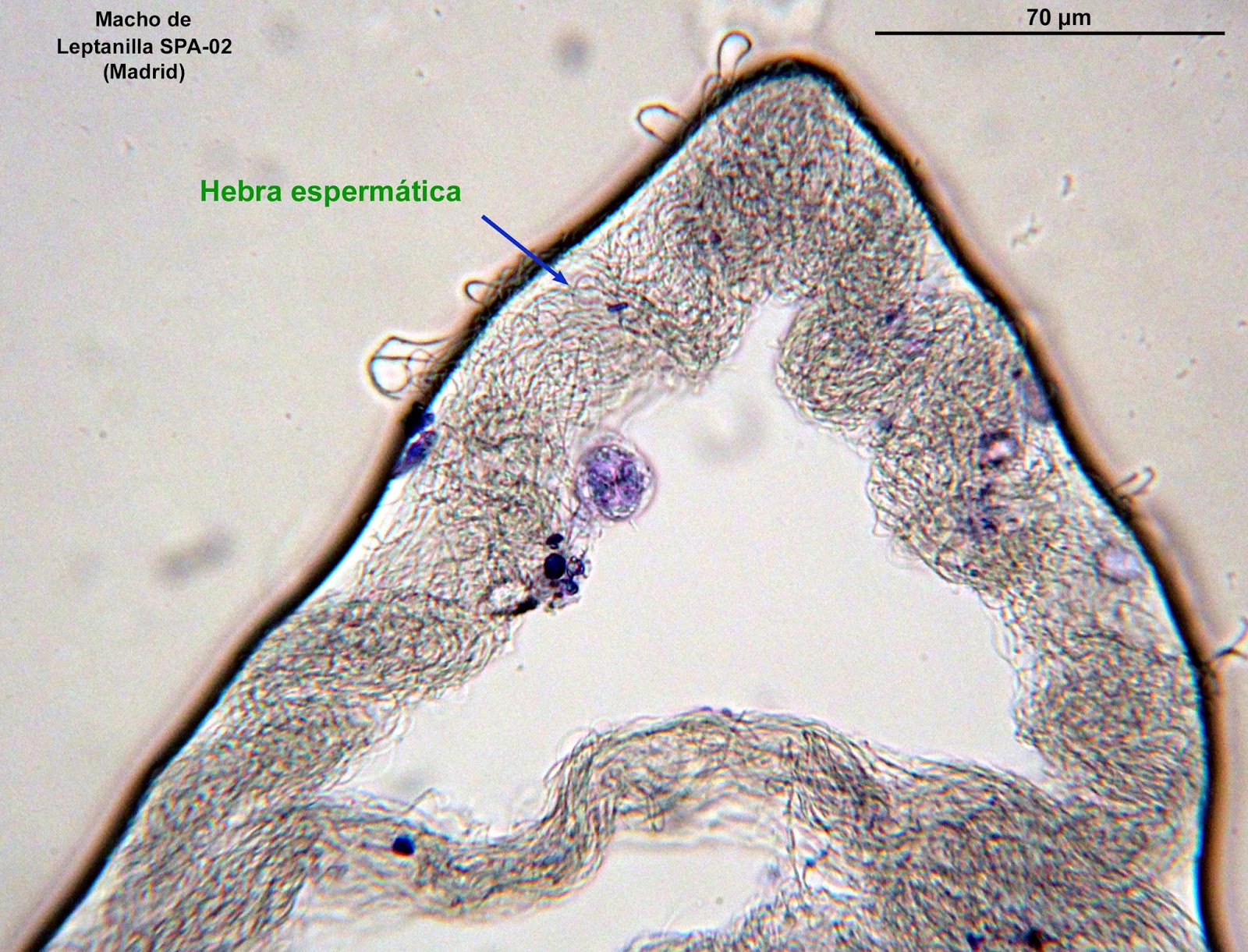
A 1000 aumentos se confirmó la naturaleza espermática de las hebras, consistente en una intrincada red de filamentos aparentemente independientes, esto es, no formando agrupaciones de espermatozoides unidos por las cabezas, como se conoce en algunos otros grupos de hormigas. Aunque no pude detectar ningún espermatozoide aislado, el seguimiento de varios extremos sueltos indica que superan las 30 micras de longitud.

Vayamos finalmente al sentido biológico de estas extrañas hebras. Mi impresión es que son el resultado meramente mecánico de los movimientos compulsivos premortem a que aludí más arriba. La segregación del esperma en forma de hilo largo vendría facilitada por su rápida y libre difusión sobre la superficie del alcohol mientras el macho se agita. Esto queda refrendado en la siguiente observación: introduje un macho vivo en una placa Petri vacía, macho que murió a las pocas horas. Pasadas varias semanas, al observar a la lupa el ejemplar seco, vi que tenía adherido en el extremo del edeago una sustancia dura y translúcida de aspecto ambarino. Se trataba del esperma eyaculado, compactado y solidificado.

En estado natural es previsible que la transferencia espermática durante la cópula se realice de manera compactada, con la hebra surgiendo de la falotrema y plegándose rápida y uniformemente a medida que penetra en la bursa copulatrix de la reina. Las singulares características de las hebras segregadas durante el estrés premortem apuntan a la posibilidad de que los machos de Leptanilla transfieran a las reinas espermatóforos genuinos.
La primera propiedad, muy llamativa, que implican estas hebras es la de la inmediata solidificación del esperma cuando es segregado al exterior, fenómeno complejo que requeriría, como en el caso de la seda de las arañas y las orugas, o en los chorros utilizados por algunos onicóforos para cazar, de una explicación molecular que dé cuenta del paso repentino de la fase líquida a la sólida.
Las hebras de los machos de Leptanilla son además notablemente resistentes y elásticas. Con dos pelos de pincel, sujetando los extremos de las hebras, he podido estirarlas y contraerlas y volverlas a estirar hasta romperlas no sin ejercer cierta fuerza. Estas propiedades podrían explicarse en parte por la trama de miles de espermatozoides filamentosos que la constituyen. Pero la consistencia de dicha trama debe darla, a modo de aglutinante, el fluido seminal que los envuelve, fluido procedente de las glándulas accesorias y de cuya composición nada sabemos.
Para medir adecuadamente la hebra de un macho de Leptanilla SPA-02, la fijé primero en formol al 4%, la estiré después sobre un portaobjetos y la incluí en bálsamo de Canadá. Medía 6.34 mm

El estudio de la ultraestructura y organización de los espermatozoides de Leptanilla requerirá del empleo de la microscopía electrónica. Estas largas hebras, no muy difíciles de obtener con las trampas adecuadas para capturar machos vivos, podrían facilitar el trabajo evitando complejas disecciones en hormigas tan pequeñas.
Leptanilla en mi barrio
Hace poco narraba mi sorpresa por el hallazgo de la hormiga amazona, Polyergus rufescens, en el jardín de unos edificios cercanos a mi casa de Madrid, en el populoso barrio de Moratalaz.
Desde hace tiempo, cuando paseamos por una de las pocas áreas no urbanizadas del barrio, cercana al cementerio de la Almudena, he comentado a mi mujer la posibilidad de que allí mismo, a 600 metros de casa, hubiera Leptanilla, uno de los géneros de hormigas más primitivos, minúsculos y difíciles de colectar. En tiempos pasados Moratalaz fue una meseta con huertas y pastos surcada por numerosos arroyos.
Junto al cauce seco de uno de esos antiguos arroyos coloqué hace tres semanas una cubeta con agua, a la que acudía cada 2 o 3 días para rellenarla. El objetivo era ver si caía algún macho de Leptanilla. En la siguiente imagen muestro la cubeta entre matojos, con el Pirulí al fondo.

Tras 20 días ha caído un macho del tipo spa-03, tipo que ya había colectado en mi habitual área de búsqueda y excavación a unos 11 km de Moratalaz.

Este hallazgo ha tenido un punto de emoción. La franja no urbanizada entre el cementerio de la Almudena y Moratalaz es una especie de isla delimitada por carreteras e innumerables edificios. La experiencia me hace sospechar que allí debe haber abundantes colonias de Leptanilla cercanas entre sí.

Quería saber si la columna utiliza el sol (y los patrones de luz polarizada) durante su avance. Cogí un trozo de cartón y le hice en el centro un pequeño agujero sobre el que coloqué la cámara de un teléfono móvil. Con dicho cartón le taparía el sol y el cielo azul al frente de la columna dePolyergus, filmando el proceso al mismo tiempo. Otra persona reflejaría la luz solar desde otro ángulo mediante un espejo.

El resultado, repetido varias veces, fue inequívoco: las obreras del frente de la columna variaban su dirección de acuerdo con el ángulo de la luz reflejada por el espejo. En el siguiente esquema muestro uno de los experimentos. La columna partió del nido con un ángulo de 90º respecto del sol. Al reflejar la luz desde otro lado, las obreras modificaron la dirección de avance para mantener el ángulo de 90º.

Este es el pequeño video del cambio de dirección de la columna al reflejarle la luz solar
https://youtu.be/InE8giJ7SZc
Suscribirse a:
Entradas (Atom)