Los Virus Por el Ing.Agr. Carlos González
El virus es un agente genético que posee una región central de ácido nucleico, ADN o ARN (genoma) y que está rodeado por una cubierta de proteína o cápside y, en algunos casos, por una envoltura lipoproteica.
Los virus contienen toda la información necesaria para su ciclo reproductor; que solamente puede ocurrir adentro de las células vivas, apoderándose de las enzimas y de la maquinaria biosintética de sus hospedadores.
Los virus difieren entre sí por el tamaño, la forma y la composición química de su genoma.
EL GENOMA
En algunos virus, el genoma se presenta segmentado en 8 moléculas de RNA de cadena simple, que se asocian con moléculas de una proteína que le confieren forma helicoidal. Los RNA genómicos asociados con la proteína reciben el nombre de nucleocápsides. Rodeando las nucleocápsides, existe una membrana lipoproteica a través de la cual emergen las glucoproteínas virales de envoltura (neuroaminidasa y hemaglutinina).
| 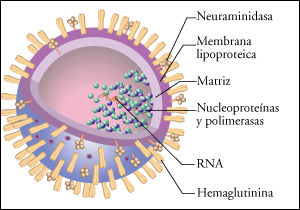 |
¿Sabía que...?
El virus de la influenza muta frecuentemente. Los cambios en su ácido nucleico alteran las proteínas de la envoltura externa y, por lo tanto, los anticuerpos previamente formados ya no lo "reconocen". Es probable que surjan nuevas cepas de virus de influenza más rápidamente que las vacunas que puedan producirse para combatirlas.
|
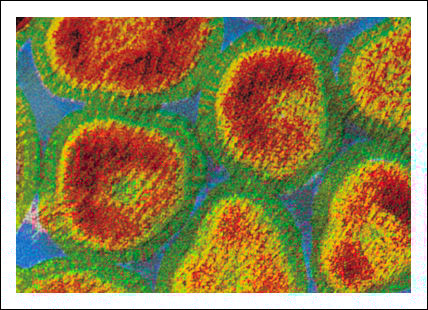 virus de la influenza(M.E.) virus de la influenza(M.E.) |
CLASIFICACIÓN
A) En las primeras épocas se tenían en cuenta los siguientes factores:
- La patogenicidad;
- El órgano o tejido atacado; y
- El tipo de transmisión.
B) En el presente, merced a la microscopía electrónica, se tienen en cuenta:
- La forma o estructura; y
- El tamaño
Tipos de estructuras:
* Helicoidal
En este tipo de estructura, los cápsides se agrupan y se ensamblan formando una hélice cerrada, en cuyo espacio medio se encuentra el genoma.
|  |
*Icosaédrica:
Cada uno de los veinte lados de esta estructura es un triángulo equilátero, compuesto por subunidades proteicas idénticas. Muchos virus están constituidos sobre este principio. Hay 252 subunidades en total. Dentro del icosaedro se encuentra el genoma viral de DNA de doble cadena.
|  |
* "T4". (bacterófagos)
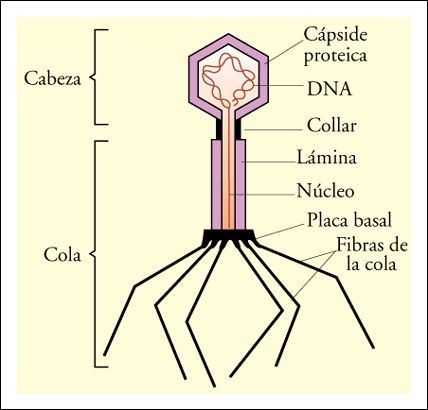 | 
Foto con microscopio electrónico
|
C) La biología molecular estudia los virus considerando que:
1- El genoma de los virus puede estar constituido por DNA o RNA de cadena simple o doble.
2- Las proteínas de la cápside pueden tomar distintas formas, que son:
a. Capas adicionales; y
b. Estructuras proteicas complejas
3- La envoltura lipídica, proveniente de la célula infectada, en la que están insertadas proteínas virales. La mayor parte de esas proteínas están glucosiladas y se denominan glucoproteínas de envoltura.
¿Sabia que...?
Las proteínas de la cápside o las proteínas de envoltura determinan la especificidad de un virus; una célula puede ser infectada por un virus si la proteína viral puede "encajar" en uno de los receptores específicos de la membrana celular de ese tipo de célula.
|
Aparentemente, todos los tipos de células, tanto procarióticas como eucarióticas, son susceptibles de infección por virus específicos capaces de establecer una interacción con sus receptores de membrana.
Los virus pueden actuar de dos formas distintas:
· Reproduciéndose en el interior de la célula infectada, utilizando todo el material y la maquinaria de la célula hospedante.
· Uniéndose al material genético de la célula en la que se aloja, produciendo cambios genéticos en ella.
Por eso se pueden considerar los virus como agentes infecciosos productores de enfermedades o como agentes genéticos que alteran el material el material hereditario de la célula huésped.
Ciclo de multiplicación de los distintos virus:
La única función que cumplen los virus y que comparten con el resto de los seres vivos es la de reproducirse (generar copias de sí mismos); para ello, necesitan utilizar la materia, la energía y la maquinaria de la célula huésped, por lo que se los denomina parásitos obligados. Como no poseen metabolismo ni organización celular, se los sitúa en el límite entre lo vivo y lo inerte.
Una vez que infectan una célula, los virus pueden desarrollar dos tipos de comportamiento: a) como agentes infecciosos, produciendo la lisis o muerte de la célula, o b) como virus atenuados o templados, que añaden material genético a la célula hospedante y, por lo tanto, resultan agentes de la variabilidad genética.
Ambos casos han sido estudiados con detalle en los virus bacteriófagos, que pueden observarse en estos dibujos esquemáticos.
| 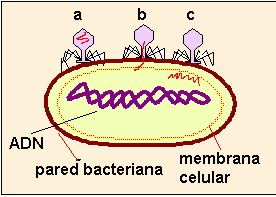 |
En los dos casos de infección el proceso empieza de esta forma:
1. Fase de fijación (a): Los virus se unen por la placa basal a la cubierta de la pared bacteriana.
2. Fase de contracción (b): La cola se contrae y el ácido nucleico del virus empieza a inyectarse.
3. Fase de penetración (c): El ácido nucleico del virus penetra en el citoplasma de la bacteria, la cubierta proteínica (cápsides) queda fuera de la célula.
A partir de este momento el proceso infeccioso puede seguir dos ciclos diferentes:
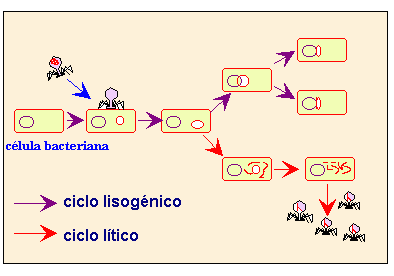 |
1. En el ciclo lítico El ADN del virus codifica todas las proteínas necesarias, el ADN bacteriano fabrica las proteínas víricas y copias de ácidos nucleicos víricos, la cabeza de la cápside, las estructuras más importantes de la cola y las fibras de la cola que se ensamblan por separado.
Después de que el DNA vírico ha sido insertado en la cabeza de la cápside, el ensamble de la cola preformada se une a ella. La adición de las fibras de la cola completa la partícula viral.
Cuando hay suficiente cantidad, los virus se liberan al medio, produciendo la muerte de la célula.
2. En el ciclo lisogénico se produce cuando el genoma del virus queda integrado en el genoma de la bacteria, no expresa sus genes y se replica junto al de la bacteria.
El virus queda en forma de profago.
Y en esta secuencia animada, puedes ver a un virus en acción...
 |
En otras, el virus intacto entra a la célula pero, una vez dentro, la cápside se desensambla por distintos mecanismos, liberando el ácido nucleico viral. Luego, el genoma viral comienza a transcribirse y a replicarse y forma nuevas partículas virales. La estrategia que utilizan los virus para multiplicarse varía de acuerdo al tipo de virus, lo que determina, a su vez, el lugar dentro de la célula en que se replica y transcribe su genoma.
Actividad biosintética del los Virus:
En los virus con genoma de ADN, el ADN del virus se replica y también se transcribe a ARN mensajero (mARN). El mARN codifica enzimas virales, proteínas de la cubierta viral y, en algunos casos, proteínas reguladoras que controlan la expresión del genoma de la célula hospedadora. El virus realiza sus actividades biosintéticas con el equipamiento de la célula hospedadora. Muchos virus usan enzimas del hospedador al igual que las codificadas por sus propios ácidos nucleicos; algunos fragmentan el ADN del hospedador y reciclan los nucleótidos para la síntesis del ADN viral. En la mayoría de los virus de ARN, el ARN viral se replica y actúa directamente como mARN. Otros en cambio, llevan en la partícula viral una enzima propia que les permite sintetizar los mARN, usando como molde el ARN genómico, ya que éste no puede funcionar como mensajero.
En otro tipo de virus de ARN, el ARN viral se transcribe a ADN a partir de éste vuelve a transcribirse luego el mARN. Este fenómeno de transcripción inversa es característico de los retrovirus, tanto de los que causan cáncer, como del virus HIV, responsable del SIDA (síndrome de inmunodeficiencia adquirida).
Las partículas virales se ensamblan dentro de la célula hospedadora. Los virus recién formados surgen como brotes en porciones de la membrana de la célula hospedadora que contienen las proteínas virales y, al hacerlo, quedan envueltos por fragmentos de ella.
Cuando se ha completado el ensamble de partículas virales, éstas se desprenden de la célula hospedadora, a menudo provocando la lisis de su membrana en el proceso. Cada nueva partícula viral es capaz de comenzar un nuevo ciclo de infección en una célula no infectada.
Evolución de los virus:
El estudio del origen y de la evolución de los virus se ve dificultado por la falta de restos fósiles. Los síntomas de enfermedades virales que conocemos actualmente pueden ser rastreados sólo hacia el comienzo de los registros de la historia humana.
Para realizar estudios comparativos, sólo disponemos de virus aislados hace no más de 80 años. Por lo tanto, para elaborar una hipótesis sobre el origen de los virus, solo podemos hacer extrapolaciones hacia atrás, basándonos en el estudio detallado de las características de los virus actuales.
Existen tres teorías principales que explicarían el origen de los virus. Una de ellas, la teoría regresiva, propone a los virus como formas degeneradas de parásitos intracelulares. Otra teoría postula que los virus se habrían originado a partir de componentes celulares normales (ADN o ARN) que habrían adquirido la capacidad de replicarse en forma autónoma y de evolucionar independientemente. La tercera teoría se relaciona con la hipótesis de un mundo prebiótico basado en ARN.
Han sido aislados e identificados otros agentes infecciosos aun más simples que los virus: los viroides (pequeñas moléculas de ARN sin proteínas asociadas) y los priones. Los viroides son el agente causal de ciertas enfermedades de las plantas y los priones transmiten enfermedades neurodegenerativas llamadas encefalopatías espongiformes. No se conocen los mecanismos por los cuales los viroides ejercen sus efectos patogénicos, pero una hipótesis sugiere que estos elementos interfieren con la regulación génica de las células infectadas.
Envoltura membranosa de los virus
La cubierta exterior de un virus puede ser una cápside proteica o una envoltura membranosa:
Inicialmente se pensó que la cubierta externa de los virus estaba formada por un sólo tipo de molécula proteica. Se creía que las infecciones víricas empezaban por la separación del cromosoma vírico (su ácido nucleico) de la cubierta proteica, y que a continuación se producía la replicación del cromosoma dentro de la célula huésped, generándose muchas copias idénticas. Después de la síntesis de nuevas copias de la envoltura proteica específica del virus sobre moléculas de RNA mensajero codificadas por el virus, podía producirse la formación de partículas víricas hijas mediante el ensamblaje espontáneo de esta cubierta proteica rodeando los cromosomas víricos hijos
Ahora se sabe que estas ideas sobre simplifican extraordinariamente la diversidad de ciclos vitales víricos que existen. Por ejemplo, la proteína de cubierta que rodea el ácido nucleico de la mayoría de los virus (la cápside) contiene más de un tipo de cadena polipeptídica, a menudo dispuesta en varias capas. En muchos virus, además, la cápside proteica está recubierta a su vez por una membrana formada por una bicapa lipídica que contiene proteínas. Muchos de estos virus recubiertos adquieren su envoltura durante el proceso de brote de la membrana plasmática . Este proceso de gemación permite a las partículas víricas abandonar la célula sin romper la membrana plasmática y, por lo tanto, sin matar a la célula.
Los virus utilizan la maquinaria de tráfico intracelular de sus células huéspedes
Todos los virus tienen una cantidad limitada de ácido nucleico en su genoma, por lo que han de parasitar procesos de la célula huésped para la mayoría de las etapas de su producción. De hecho, debido a que habitualmente los productos víricos se sintetizan en grandes cantidades durante la infección y debido también a que durante su ciclo vital los virus siguen una rota secuencial a través de los compartimientos de la célula huésped, las células infectadas por virus se han utilizado como importantes modelos para trazar las rutas de
| 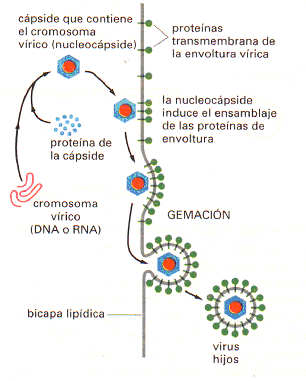 Fig. 01 |
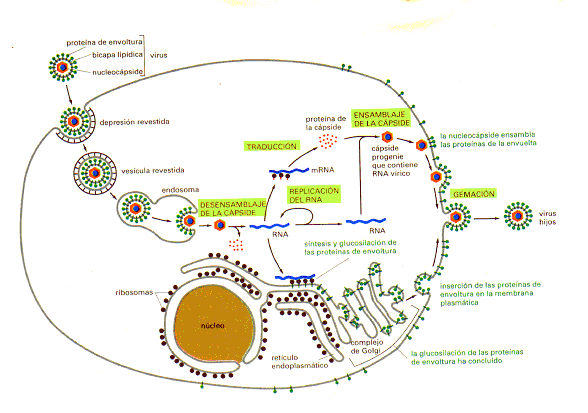
Fig. 02 Ciclo vital del virus Semliki forest. El virus parasita la célula huésped para la mayor parte de sus procesos biosintéticos.
|
transporte intracelular y para estudiar qué reacciones biosintéticas esenciales están comparomentalizadas en las células eucariotas.
Los virus recubiertos que afectan a las células animales, en los cuales el genoma está incluido en una membrana de bicapa lipídica, han utilizado la compartimentalización de la célula hasta un grado especialmente preciso. Seguir el ciclo vital de un virus recubierto es hacer un "tour" a través de la célula. Un ejemplo bien estudiado es el virus Semliki forest, que consiste en un genoma de cadena sencilla de ARN rodeado por una cápside formado por una envoltura eicosaédrica (20 caras) dispuesta de forma regular y compuesta por muchas copias de una proteína (denominada proteína C). La nucleocápside (genoma + cápside) está rodeada por una bicapa lipídica situada muy cerca, que contiene únicamente tres tipos de cadenas polipeptídicas, codificadas por el ARN vírico. Estas proteínas de envoltura forman heterotrímeros situados en la bicapa lipídica y que interactúan con la proteína C de la nucleocápside, uniendo la membrana con la nucleocápside. Las zonas glucosiladas de las proteínas de envoltura siempre están situadas en el exterior de la hicapa lipídica, y cada trímero forma una "espina " que al
|
microscopio electrónico se ve sobresaliendo de la superficie del virus.
La infección se inicia cuando una proteína de envoltura del virus se une a una proteína de una célula normal que actúa como su receptor en la membrana plasmática de la célula huésped. Entonces el virus utiliza el proceso endocítico normal de la célula para entrar en ella mediante endocitosis mediada por receptor, y es liberado a los endosomas vecinos. Sin embargo, en lugar de ser transferido desde los endosomas a los lisosomas, el virus se escapa de los endosomas gracias a las propiedades especiales de una de sus proteínas de envoltura. Al pH ácido del endosoma esta proteína hace que la envoltura vírica se fusione con la membrana del endosoma, liberando la nucleocápside desnuda en el citoplasma. La nucleocápside pierde su cubierta en el citoplasma, liberando el ARN vírico, que entonces es traducido por los ribosomas de la célula huésped produciendo una ARN polimerasa codificada por el virus. Esta enzima, a su vez, produce muchas copias del ARN vírico, algunas de las cuales actúan como moléculas de ARNm dirigiendo la síntesis de proteínas estructurales del virus-la proteína C de la cápside y las tres proteínas de la envoltura.
Las proteínas de la cápside y de la envoltura, recién sintetizadas, siguen vías separadas a través del citoplasma. Las proteínas de la envoltura, como las proteínas de la membrana plasmática de la célula huésped, son sintetizadas en los ribosomas que se hallan unidos al ER rugoso; por el contrario, la proteína de la cápside, como las proteínas citosólicas de la célula, se sintetiza por ribosomas que no se hallan urudos a membrana. Las proteínas recién sintetizadas de la cápside se unen al RNA vírico recientemente replicado, formando nuevas nucleocápsides. Las proteínas de la envoltura, por el contrario, son insertadas en la membrana del ER, donde son glucosiladas, transportadas hasta el complejo de Golgi y liberadas a la membrana plasmática .
Finalmente las nucleocápsides víricas y las proteínas de la envoltura se encuentran en la membrana plasmática. Como resultado de una interacción específica con un grupo de proteínas de envoltura, la nucleocápside forma una gema cuya envoltura contiene proteínas de envoltura embebidas en lípidos de la célula huésped. Por último, la gema se libera, de forma que aparece un nuevo virus independiente de la célula. E1 agrupamiento de las proteínas de la envoltura mientras se ensamblan alrededor de la nucleocápside durante la gemación vírica hace que las proteínas plasmáticas de la célula huésped queden excluidas de la partícula vírica final.
No hay comentarios:
Publicar un comentario